Advancing Cancer Diagnostics, Improving Lives
La villine, ainsi que les protéines apparentées, la gelsoline, la fragmine et la séverine, régulent l'architecture et l'assemblage de l'actine.
La villine est unique parmi ces protéines par sa capacité à réticuler les filaments d'actine en faisceaux, un processus qui ne s'observe qu'à de faibles concentrations en Ca2+.
La villine est constituée de trois domaines. Les deux premiers domaines sont homologues, et le troisième domaine est appelé domaine d'en-tête. Cette région d'en-tête se trouve à l'extrémité C-terminale.
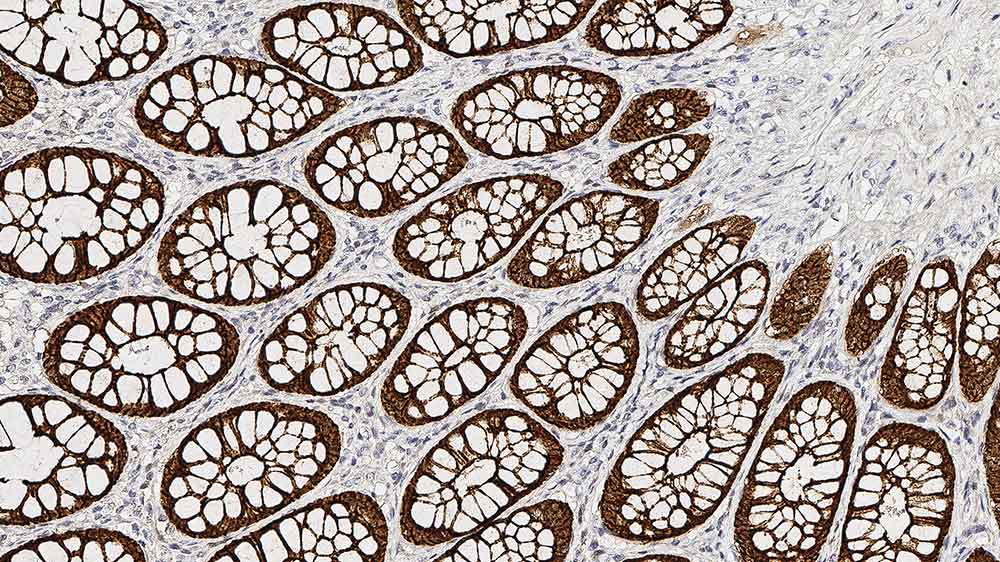
La villine est principalement produite par les cellules épithéliales qui développent une bordure en brosse.
Les cellules productrices de villine ont été observée soit à la surface des cellules épithéliales des muqueuses de l'intestin et de la vésicule biliaire, ou à la surface des cellules épithéliales des tubules proximaux du rein et des cônes efférents des testicules.
On a cependant également observé la villine dans certains épithéliums sans bordure en brosse, mais dérivés de l'intestin embryonnaire, comme les cellules des canaux du pancréas exocrine et les cellules biliaires du foie. Dans ces types cellulaires, la villine se concentre dans le cytoplasme apical.
Les cellules épithéliales de la muqueuse intestinale sont continuellement renouvelées, ce qui implique la migration de ces types cellulaires des cryptes intestinales vers les extrémités des villosités, migration accompagnée de l'acquisition graduelle de leur phénotype différencié. La production maximale de villine se produit à la base des villosités.
La villine possède donc une expression spécifique au tissu, limitée à certains épithéliums et à leur domaine apical, et indique dès lors leur polarité.
La perte morphologique de polarité des cellules épithéliales du côlon est un des indicateurs les plus significatifs des dysplasies et des néoplasies.